Notes on Draper's Article on Behe's Design Argument, Part 3: Are Behe's Examples Really Irreducibly Complex?
We've been discussing Paul Draper's criticisms of Behe's design argument in Draper's 2002 article, "Irreducible Complexity and Darwinian Gradualism: A Reply to Michael J. Behe".[1] To briefly review,
recall that the article focuses on stage one of Behe's two-stage design argument, which argues that certain biochemical structures cannot have arisen via gradualistic Darwinian processes. The argument of this stage crucially relies on his notion of irreducible complexity, where this is defined as a system "composed of several well-matched, interacting parts that contribute to the basic function, wherein removal of any of the parts causes the system to effectively cease functioning". [2] With this notion in hand, Behe argues that there are irreducibly very complex biochemical systems, and that these systems can't plausibly be explained in terms of gradualistic evolutionary processes. And the reason is that evolution can only create systems via direct and indirect evolutionary pathways. But evolution can create no irreducibly complex system via a direct evolutionary pathway. And while evolution can create simple irreducibly complex systems via indirect pathways, and reducibly complex systems via direct and indirect evolutionary pathways, the odds are overwhelmingly against creating irreducibly very complex systems via indirect pathways.
That's the argument laid out in Part I of Behe's book. In part II, Behe attempts to support the key premise that some biochemical systems are irreducibly (very) complex. Toward that end, he gives seven examples of such systems: (i) the bacterial flagellum, (ii) the cilium, (iii) the vertebrate blood-clotting system, (iv) certain cellular transport systems, and three subsystems of our immune system: (v) the clonal selection system (vi) the antibody-diversity system, and (vii) the complement system. Given this, and given his argument, there are at least three ways to criticize his argument directly:
(1) Undercut or rebut the claim that his example systems are irreducibly complex (or at least irreducibly very complex)
(2) Undercut or rebut the claim that irreducibly complex systems can't be created via indirect evolutionary pathways
(3) Undercut or rebut the claim that irreducibly complex systems can't be created via direct evolutionary pathways
Draper argues that Behe's argument falls prey to all three types of criticism. In this installment, I'll cover most of his discussion of type-(1) criticisms.
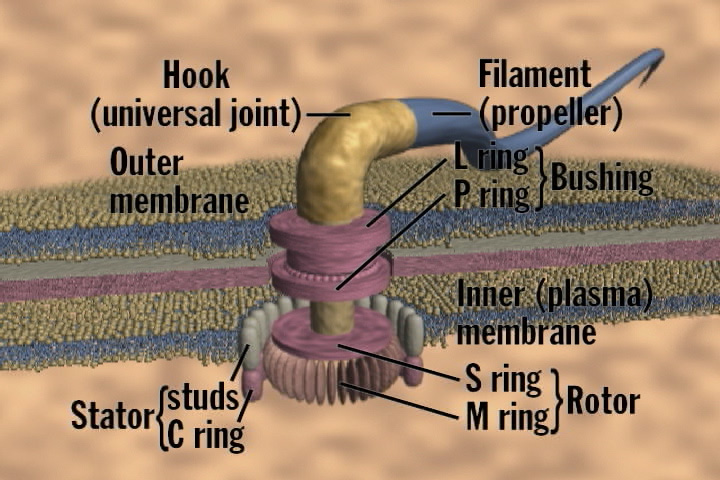
First, Draper points to the work of others to offer rebutting defeaters for a number of Behe's candidates of irreducibly complex systems. So, for example, consider Behe's cilium example. Behe argues that cillia require eleven microtubules to function: two central microtubules surrounded by nine encasing microtubules. But Draper refers to Ken Miller's point that some organisms have cilia with three microtubules and no inner microtubules. Therefore, since cilia don't require all eleven parts to function, Behe's cillium example fails to satisfy his own account of irreducible complexity. A similar problem plagues Behe's flagellum example. Behe asserts that the bacterial flagellum requires at least 40 parts to function. But Draper refers to biochemist David W. Ussery's point that some forms of bacteria have flagella that only require 33 parts to function. But if so, then since the bacterial flagellum doesn't require all 40 parts to perform its function, then by Behe's definition, it isn't irreducibly complex.[4] Draper points out that the same criticism applies to Behe's immune system and cellular transport examples.
Second, Draper points out that Behe fails to show that his own example systems are irreducibly complex. On p. 42 of Darwin's Black Box, Behe states his two-step method of testing and demonstrating whether a system is irreducibly complex:
Step 1: Find the system's function, and identify all the components that contribute to that function.
Step 2: Determine whether all of the system's components are required for it to perform the function.
But when it comes time to argue that his example systems are irreducibly complex (in Part II of the book), he fails to follow (at least) Step 2 for any of his example systems. Instead, he typically picks a proper subset of a system's total components, and argues that the system can't function without them. So, for example, in his discussion of the cilium on p. 73, he says that it has "dozens or even hundreds" of parts involved in its function. However, instead of explaining how each of these parts is necessary for the function of the system, he picks four of them, discusses their essentiality for performing the paddling function, and then fleetingly asserts that probably many other of its parts are essential as well [3]. As mentioned above, and as Draper points out, Behe continues to ignore Step 2 in his presentations of the remaining five example systems as well. The result is that a crucial premise in the main argument of Behe's book -- that there are irreducibly very complex biochemical systems -- is left unargued for.
Draper nicely sums up the problems with Behe's claim that some biochemical structures are irreducibly (very) complex: "The bottom line is that Behe doesn't deliver in the second part of his book what he promised in the first part...The systems upon which he bases his case contain parts that contribute to the system's function, yet either are not essential for that function or at least have not been shown by Behe to be so."[5]
As Draper points out, however, Behe has come up with a reply to this criticism. We'll look at it in the next installment.
----------------------------------------------------------------
Notes
[1]Faith and Philosophy 19:1, pp. 3-21.
[2] Behe, Michael J. Darwin's Black Box: The Biochemical Challenge to Evolution (New York: The Free Press, 1996), p. 39.
[3] p. 73
[4] I should note that Kenneth Miller has gone farther on this point by showing that the parts of the flagellum system, while irreducibly complex qua propeller, has plausible evolutionary precursors that performed different functions. See this YouTube clip for a sketch of his point.
A somewhat technical quibble: There seems to be some ambiguity with respect to Behe's notion of irreducible complexity, leaving room for two interpretations of a clause in his notion of irreducible complexity (the one about loss of function with the removal of a part):
Interpretation 1: If any part of the system were removed, it would cease to have any useful biological function for the organism.
Interpretation 2: If any part of the system were removed, it would cease to have the function it currently has (although not necessarily some other).
Now Miller's point in the clip is that, contrary to what Behe has argued, the bacterial flagellum example isn't irreducibly complex. However, in light of the two interpretations above, we see that whether Miller's example in the clip supports that point depends on whether interpretation 1 is correct; If it's not, Miller's point is false.
I say that this is a something of a quibble because the substance of Miller's point poses a problem for Behe's account even if he has misinterpreted Behe's notion of irreducible complexity (although I don't think he has). For Miller's example shows a plausible evolutionary precursor to the bacterial flagellum. And given this, MIller's example poses a problem for Behe's use of the bacterial flagellum in his design argument on either intperpretation: On interpretation 1, Miller's example undercuts Behe's claim that the bacterial flagellum is irreducibly complex. And on interpretation 2, Miller's example undercuts Behe's claim that irreducibly very complex systems (such as the bacterial flagellum) cannot be created via an evolutionary pathway.
[5] Draper, ibid, p. 10.
recall that the article focuses on stage one of Behe's two-stage design argument, which argues that certain biochemical structures cannot have arisen via gradualistic Darwinian processes. The argument of this stage crucially relies on his notion of irreducible complexity, where this is defined as a system "composed of several well-matched, interacting parts that contribute to the basic function, wherein removal of any of the parts causes the system to effectively cease functioning". [2] With this notion in hand, Behe argues that there are irreducibly very complex biochemical systems, and that these systems can't plausibly be explained in terms of gradualistic evolutionary processes. And the reason is that evolution can only create systems via direct and indirect evolutionary pathways. But evolution can create no irreducibly complex system via a direct evolutionary pathway. And while evolution can create simple irreducibly complex systems via indirect pathways, and reducibly complex systems via direct and indirect evolutionary pathways, the odds are overwhelmingly against creating irreducibly very complex systems via indirect pathways.
That's the argument laid out in Part I of Behe's book. In part II, Behe attempts to support the key premise that some biochemical systems are irreducibly (very) complex. Toward that end, he gives seven examples of such systems: (i) the bacterial flagellum, (ii) the cilium, (iii) the vertebrate blood-clotting system, (iv) certain cellular transport systems, and three subsystems of our immune system: (v) the clonal selection system (vi) the antibody-diversity system, and (vii) the complement system. Given this, and given his argument, there are at least three ways to criticize his argument directly:
(1) Undercut or rebut the claim that his example systems are irreducibly complex (or at least irreducibly very complex)
(2) Undercut or rebut the claim that irreducibly complex systems can't be created via indirect evolutionary pathways
(3) Undercut or rebut the claim that irreducibly complex systems can't be created via direct evolutionary pathways
Draper argues that Behe's argument falls prey to all three types of criticism. In this installment, I'll cover most of his discussion of type-(1) criticisms.
First, Draper points to the work of others to offer rebutting defeaters for a number of Behe's candidates of irreducibly complex systems. So, for example, consider Behe's cilium example. Behe argues that cillia require eleven microtubules to function: two central microtubules surrounded by nine encasing microtubules. But Draper refers to Ken Miller's point that some organisms have cilia with three microtubules and no inner microtubules. Therefore, since cilia don't require all eleven parts to function, Behe's cillium example fails to satisfy his own account of irreducible complexity. A similar problem plagues Behe's flagellum example. Behe asserts that the bacterial flagellum requires at least 40 parts to function. But Draper refers to biochemist David W. Ussery's point that some forms of bacteria have flagella that only require 33 parts to function. But if so, then since the bacterial flagellum doesn't require all 40 parts to perform its function, then by Behe's definition, it isn't irreducibly complex.[4] Draper points out that the same criticism applies to Behe's immune system and cellular transport examples.
{kind=link}
Second, Draper points out that Behe fails to show that his own example systems are irreducibly complex. On p. 42 of Darwin's Black Box, Behe states his two-step method of testing and demonstrating whether a system is irreducibly complex:
Step 1: Find the system's function, and identify all the components that contribute to that function.
Step 2: Determine whether all of the system's components are required for it to perform the function.
But when it comes time to argue that his example systems are irreducibly complex (in Part II of the book), he fails to follow (at least) Step 2 for any of his example systems. Instead, he typically picks a proper subset of a system's total components, and argues that the system can't function without them. So, for example, in his discussion of the cilium on p. 73, he says that it has "dozens or even hundreds" of parts involved in its function. However, instead of explaining how each of these parts is necessary for the function of the system, he picks four of them, discusses their essentiality for performing the paddling function, and then fleetingly asserts that probably many other of its parts are essential as well [3]. As mentioned above, and as Draper points out, Behe continues to ignore Step 2 in his presentations of the remaining five example systems as well. The result is that a crucial premise in the main argument of Behe's book -- that there are irreducibly very complex biochemical systems -- is left unargued for.
Draper nicely sums up the problems with Behe's claim that some biochemical structures are irreducibly (very) complex: "The bottom line is that Behe doesn't deliver in the second part of his book what he promised in the first part...The systems upon which he bases his case contain parts that contribute to the system's function, yet either are not essential for that function or at least have not been shown by Behe to be so."[5]
As Draper points out, however, Behe has come up with a reply to this criticism. We'll look at it in the next installment.
----------------------------------------------------------------
Notes
[1]Faith and Philosophy 19:1, pp. 3-21.
[2] Behe, Michael J. Darwin's Black Box: The Biochemical Challenge to Evolution (New York: The Free Press, 1996), p. 39.
[3] p. 73
[4] I should note that Kenneth Miller has gone farther on this point by showing that the parts of the flagellum system, while irreducibly complex qua propeller, has plausible evolutionary precursors that performed different functions. See this YouTube clip for a sketch of his point.
A somewhat technical quibble: There seems to be some ambiguity with respect to Behe's notion of irreducible complexity, leaving room for two interpretations of a clause in his notion of irreducible complexity (the one about loss of function with the removal of a part):
Interpretation 1: If any part of the system were removed, it would cease to have any useful biological function for the organism.
Interpretation 2: If any part of the system were removed, it would cease to have the function it currently has (although not necessarily some other).
Now Miller's point in the clip is that, contrary to what Behe has argued, the bacterial flagellum example isn't irreducibly complex. However, in light of the two interpretations above, we see that whether Miller's example in the clip supports that point depends on whether interpretation 1 is correct; If it's not, Miller's point is false.
I say that this is a something of a quibble because the substance of Miller's point poses a problem for Behe's account even if he has misinterpreted Behe's notion of irreducible complexity (although I don't think he has). For Miller's example shows a plausible evolutionary precursor to the bacterial flagellum. And given this, MIller's example poses a problem for Behe's use of the bacterial flagellum in his design argument on either intperpretation: On interpretation 1, Miller's example undercuts Behe's claim that the bacterial flagellum is irreducibly complex. And on interpretation 2, Miller's example undercuts Behe's claim that irreducibly very complex systems (such as the bacterial flagellum) cannot be created via an evolutionary pathway.
[5] Draper, ibid, p. 10.